从双孢蘑菇(Agaricusbisporus)株中排除攻击性不相容性状的方法以及相关菌株和株系与流程
本发明一般地涉及从双孢蘑菇(agaricus bisporus)株中排除攻击性不相容(aggressive incompatibility)性状(也称为ai性状)的方法。还公开了与该方法相关的多种新菌株和株系。
背景技术:
1、本发明在如美国专利no.10,440,930中所公开的,由sylvan america,inc.开发的命名为j15987(nrrl保藏号67646)的较早的白色杂交蘑菇株的发布之后,该专利的公开内容作为参考并入本文。株j15987比目前主要的白色栽培品种具有更高的产量并且生产更高质量的蘑菇,这意味着这些蘑菇与从商业可接受的命名为a-15的株所获得的标准商品化白色蘑菇相比,具有更厚的菌盖肉质,并且具有更少的红色。在株j15987的早期测试中,消费者对产量和质量的强大组合非常满意,这使得这些蘑菇和该株最初在商业上非常受欢迎。然而,出现了一个问题。据发现株j15987包含一种前所未有的表型或性状,其被称为“攻击性不相容性”,当这些农场也使用株a-15或类似株时,这会导致在蘑菇农场层面出现问题。因此,需要一种将从类似于j15987的白色蘑菇株中排除“攻击性不相容性”(如以下所定义的)的表型或性状的方法,并且开发保留或另外提供株j15987的良好性状(如高产量和/或高质量),同时排除攻击性不相容性(ai)性状的这些类似的白色蘑菇株。
2、食用蘑菇,双孢蘑菇(lange)imbach双孢品种(agaricus bisporus(lange)imbachvar.bisporus)是担子菌类真菌,在全世界种植。在欧洲和美洲,它是最广泛种植的蘑菇物种。2020-21年,美国每年的作物价值为约10亿美元(国家农业统计局/usda数据)。
3、以与其它微生物类似的方式在无菌培养基上制备、维持、繁殖和储存伞菌属(agaricus)培养物。在无菌区内使用无菌工具和无菌技术来操作用于多种目的的纯培养物,包括孢子萌发、制备新的杂交体和制备接种物。商品化培养基是“菌种体”,其可以具有一些不同配方,最常见的是无菌小米或黑麦。菌种体的生产通常是大规模的。例如,1升纯伞菌属接种物可用于14,0000升灭菌的菌种体。最终用户,即蘑菇农场接受位于无菌包装内的纯菌种体培养物。
4、在蘑菇农场专门建造的构造内商业种植蘑菇。尽管在方法上有多种变化,但以下描述了典型的方法。堆肥是从废弃的木质纤维素材料,如小麦秸秆制备的,添加了含氮材料,完成并转移至特定条件下(有氧,温度在40和50℃之间)进行巴氏杀菌,借此产生有利于伞菌属定殖的基质。加入蘑菇菌种体,并在受控温度下在基质中定殖11至17天。一旦定殖或“菌种体运行(菌种体扩散,spawn run)”完成,则在堆肥表面施加无营养的饱和泥炭层(用甜菜石灰将ph调节至8至8.5之间)至约5cm的层。另外,可以将覆盖菌种体(casing spawn)添加至泥炭或覆盖层中以加速菌丝体生长。一旦发生定殖,在称为“出菇潮”的过程中调节环境条件,其中空气温度降至约18-19℃,并且co2降至约1300ppm。在施加覆土(覆盖土壤)后的第13至18天,菇房内将出菇。在3至4天的时间段内收获蘑菇。在移除和替换蘑菇农场上的堆肥之前,生产和收获额外的生产出菇潮(flush)或爆发(break)。
5、在美国,2020-21年总收获量的72%是白色伞菌属,其余是棕色伞菌属品种。同期,美国蘑菇作物的总价值超过10亿美元。在其它地区,人们对白色蘑菇有更强烈的偏好。在美国和其它地方,对于白色蘑菇,市场对于表型性状,如大小、形状、白度、硬度以及相关性状,如延长的货架期的要求是狭窄且精确的。因此,新的白色伞菌属菌株应符合一套特定的市场要求。可以基于一些性状来分离菌株,例如,菌盖形状(例如,菌盖圆度、菌肉厚度、菌柄粗度)、颜色(白度)、密度/硬度(牢固度,firmness)和单重(piece weight)。新的白色蘑菇株的商品化目标的一些实例是提高的作物产量、改变的整个生产出菇潮中的产量分布、抗病性、抗虫性、改善的货架期、减少的压伤、易于管理、对机械收获的适用性、对应激源(例如,温度或基质变化)的差异反应、季节影响、农场实践和菌株不相容性。还可以使用dna标志物技术,如单核苷酸多态性(snp),将菌株彼此区分和分离。菌株可以具有不同的祖先,这将直接反映基因型和表型。
6、大约在1980年,荷兰霍斯特(horst)的一个实验室培育出了最早两个真正的双孢蘑菇(a.bisporus)杂交株。这两个被称为u1和u3的“horst”菌株是由两组已有的白色菌株,即预杂交的光滑白色株(phsw)和预杂交的灰白色株(phow)交配所产生的密切相关的杂交体,如m.imbernon等人,mycologia,88(5),749-761(1996)中所述,该文献作为参考并入本文。随着时间的推移,杂交株u1成为市场上主要的白色基因型。
7、通过使培养物去异核化来回收株u1的亲代同核体。phsw亲代是h39,而phow亲代是h97。a.sonnenberg将h97以保藏号10389保藏在美国堪萨斯州的真菌遗传学保藏中心(fungal genetics stock center),并且还以保藏号mya-4626保藏在美国模式培养物保藏所。联合基因组计划(california,usa)对h97基因组进行了测序并将其置于公共领域,并且该基因组已被证明作为单核苷酸多态性(snp)检测的参考而非常有价值(参见下文)。
8、株u1是目前在北美洲和欧洲培养的所有白色菌株的祖代。多种蘑菇株,如a-15和s-130符合u1的实质衍生品种(edv)标准,它们是从原始(即初始)菌株的孢子或组织发育而来的。通过体细胞选择或孢子培养,或如以下所讨论的“实质衍生”的相关方法,从单一祖代所开发的菌株组群被称为衍生谱系组。除了相对较小的遗传差异外,从u1衍生谱系组内所开发的所有白色菌株与原始菌株u1共有单一的复合n+n异核基因型。除了中国明显不同之外,现代白色蘑菇栽培是单一栽培(monoculture)。
9、双孢蘑菇具有被称为同宗异宗配合(amphithallism)的生殖综合征,其中两个不同生命周期同时进行。与其它真菌中一样,生殖繁殖体是孢子。伞菌属在被称为担子的厚孢子囊(meiosporangium)上通过减数分裂产生孢子。在第一个生命周期中,双孢蘑菇孢子分别接受单个单倍体减数分裂后的核;这些孢子能够交配,但不能产生蘑菇。这些单倍体孢子萌发产生同核子代或系,其可与其它相容(亲和)的同核体交配以产生有能力产生蘑菇的新型杂交异核体。异核体通常显示出比同核体差得多的交配能力。这种生命周期被称为异宗配合(heteromixis),其类似于远交。该生命周期起作用,但通常在双孢蘑菇双孢品种(agaricus bisporus var.bisporus)菌株中不占优势。
10、第二生命周期被称为内源杂交(intramixis),其类似于近亲繁殖的形式,在大多数双孢蘑菇双孢品种菌株中占优势。大多数孢子接受两个减数分裂后的核,并且大多数这些核对由非姊妹核对(nsnp)组成,其在包括mat基因座的大多数或所有着丝粒连锁基因座处具有异等位基因基因型。mat基因型决定了这些后代由此而来的异核表型,这些子代具有繁殖能力并且可以生产蘑菇作物。不寻常地,在真核生物中,在双孢蘑菇的减数分裂后的子代中观察到发生了相对较少的染色体交叉(chromosomal crossing-over);根据经验,在异核菌株的异核内源杂交(intramictic)子代中,很少有异等位性(类似于杂合性)丢失。
11、在这两个生命周期中,孢子是作为蘑菇的一部分的细胞;该细胞是始终作为其单一亲代具有蘑菇和产生蘑菇的培养物的繁殖体。
12、就孢子而言,活孢子是处于休眠状态的异核体或同核体。孢子是蘑菇生物(有机体)培养物的一部分;它们只掺入产生它们的单一培养物(为方便起见,通常称为“亲代”)的基因材料(遗传物质)。培养物的其它部分包括菌盖、菌柄、菌褶、细胞(定义为掺入核、线粒体、细胞质、原生质体、rna、dna、蛋白质、细胞膜和细胞壁(包括交叉壁)的菌丝室(菌丝区室,hyphal compartment))、菌丝和菌丝体。可以在无菌材料上无菌收集孢子,以多种稀释度在无菌水中混悬,并铺板到无菌琼脂生长培养基上,以产生萌发的孢子和掺入孢子内的培养物。优选的技术将在封闭的培养皿内具有活的伞菌属培养物,其可以通过挥发性信息素的扩散刺激孢子萌发。萌发的孢子可以在显微镜下使用无菌显微工具,如钢针分离到新鲜的营养琼脂平板上。使用该方法,可以获得包含孢子的菌株的异核和同核子代,以及掺入到所述菌株的孢子内的培养物。
13、通过异宗配合的新型杂交品种的开发包括两个相容(亲和)的培养物的受控物理结合和交配以获得新型异核体培养物。同核体(=“系”)是用于进行交配的优选起始培养物,因为它们具有与其它培养物接合并实现胞质融合的最大能力。异核体也可以置于物理接触,但导致成功形成新型异核体的交配概率不合理地低。相容性(亲和性)由mat基因座处的基因型决定;具有相同mat等位基因的两个同核体在接合后不能建立异核体,因此同核体相容性代表遗传相异性。在确定的交配程序中,获得同核系,并以预定的成对组合相结合。在一种方法中,可以将同核体对紧密靠近地放置在培养皿中的营养琼脂培养基的表面上并使其生长在一起(处于物理结合),此时两种培养物之间发生点接合(point anastomoses)。成功的结果是提供异核体的交配。通过从培养皿的融合区转移菌丝体,可以获得新型杂交异核体。
14、出于两种原因,相对于u1衍生的谱系群,显然需要商业上可接受的具有不同基因型的白色双孢蘑菇株。首先,已知与u1衍生的谱系群的菌株营养不相容的菌株延缓病毒病在菌株之间的传播,并且已知这些疾病是商品化蘑菇产业中作物损失的主要原因。其次,众所周知,当农业产业广泛依赖于具有单一遗传谱系的作物时(如世界上大多数地方的白色双孢蘑菇),在整个设施或整个产业范围内发生灾难性作物歉收的风险就会增加。因此,从粮食安全和风险管理的角度来看,非常期望同时提供基因多样化和商业上可接受的性能和作物特征两者。
15、如以上所提及的,本发明是杂交株j15987(nrrl保藏号67646)的商业发布的直接结果,该杂交株是白色杂交株(美国专利no.10,440,930)。商品化前测试表明,与来自株a-15的其它白色栽培种相比,株j15987具有更厚的菌盖肉质和更粗的菌柄。另外,使用minolta色度计对j15987进行的颜色分析表明收获后蘑菇盖上的红色较少。商品化潜力被认为是重要的,并且该菌株在专利申请后直接在美国种植。它成功符合了所有原始作物的生产标准。然而,出现了一个显著的问题。据观察,在一些蘑姑种植“屋”(或房间)中,特别是在生产来自株a-15的蘑菇的地方,在堆肥和覆盖层两者内,都有大面积没有蘑菇生长的堆肥。很明显,这种观察现象是不正常且不寻常的。
16、将培养物返回到测试设施,在此更详细地研究了株j15987和其它白色栽培种之间的可疑拮抗相互作用。株-株相互作用的遗传学是由自身/非自身识别系统控制的,这也被称为营养不相容性(vegetative incompatibility),或者称为异核体不相容性。据发现株j15987在与其它u1家族实质衍生品种(edv),如a-15接触时具有非常具攻击性的(强烈的,aggressive)、不寻常的拮抗反应。当菌种体以低至99% a-15比1%j15987的比例混合时,伞菌属菌丝体迅速死亡,并且a-15基因型最终被j15987取代。这种相互作用被称为“攻击性不相容性(aggressive incompatibility)”,并将这种相互作用缩写为“ai”,所述表型被定义为“ai”性状。需要明确的是,包括双孢蘑菇的不相关的担子菌株通常表现出不相容性反应,这已被用于病毒控制,但ai性状是更常见现象的更具攻击性、更极端且更有问题的表现。据推测,这种性状受遗传控制,正如通常对担子菌中的营养不相容性所理解的情况一样。
17、蘑菇农场上的常规混合设备倾向于以使得不同菌株的接种基质之间可以发生一定程度混合的方式运行。这些事件通常较小,且具有适度的负面影响。因为这些区域很小,它们对总产量的影响最低,并且传统上是能容忍的。然而,更大且更极端的ai相互作用的影响过大以至于在商业上不能容忍,并且消费者(客户)的评价是在株a-15也在培养的农场操作中,无法有利地处理株j15987。因此,需要具有株j15987的积极属性同时还缺少ai性状的一种或多种商业上期望的菌株。
技术实现思路
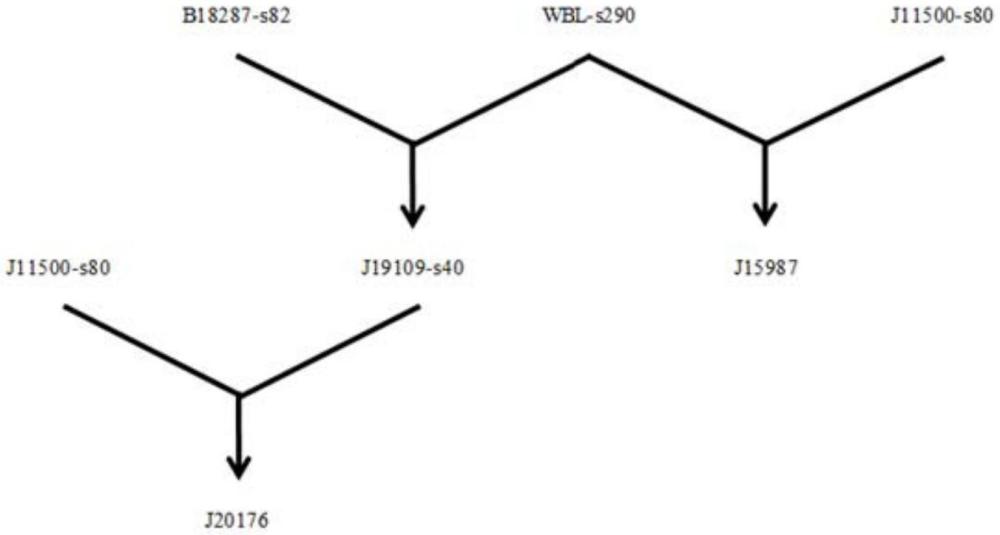
1、为了理解和克服(排除)导致株j15987中ai性状表现的潜在遗传基础,开发了一系列交配并分析了所产生的杂交体。在以下方案i中描述了交配方案。本质上,野生变异系(即b18287-s82,nrrl保藏号68168)与株j15987的同核亲代之一(即wbl-s290,nrrl保藏号68167)组合以产生新的f1杂交株(即j19109,nrrl保藏号68163)。然后,将来自该杂交株的同核体(例如,系j19109-s40,nrrl保藏号68165)与株j15987的另一同核亲代(即j11500-s80,nrrl保藏号68164)交配以提供新的f2杂交株(例如,j20176,nrrl保藏号68166)。与f2杂交株对抗(confrontation)以证实ai性状的存在或不存在。基于这些对抗,很明显交配方案是成功的,并且大多数新的杂交(体)都没有ai性状。遗传模式允许进一步开发ai性状的遗传控制/排除模型。
2、方案1
3、
4、首先揭示了:从由两个同核体(即,w01-s1和so76-s12b)交配所产生的杂交株获得的同核体(系b18287-s82)是必需的。该同核系b18287-s82是从20世纪90年代由r.kerrigan在北美洲收集的野生株w01和预杂交灰白色株so76所获得的亲代同核体开发而来的。dna测试表明w01是真正的野生株,而不是逃逸的商业株。此外,w01与目前的白色和棕色菌株具有非常类似的生长要求。
5、因此,本发明的一个方面一般地涉及命名为b18287-s82的双孢蘑菇的新的独特的同核系,以及使用命名为b18287-s82的株系的方法。在至少一个实施方式中,提供了包含双孢蘑菇系b18278-s82的至少一组染色体的培养物,其中所述至少一组(set)染色体包含下表i中列出的序列表征的标志物基因座处的系b18278-s82的所有等位基因。如本文所公开的,双孢蘑菇系b18287-s82的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815north university street,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68168。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。以适用的专利法要求的为准,在提交专利申请或颁发专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
6、在一个实施方式中,可以提供系b18287-s82的实质衍生品种(基本衍生品种,essentially derived variety)(edv)。该edv可以包括衍生自初始培养物的培养物,其中所述初始培养物是系b18287-s82的培养物,从而使得其基因组或基因型的至少75%存在于系b18287-s82的初始培养物的基因组或基因型中。在其它实施方式中,其基因组或基因型的至少90%或95%或98%或99%存在于系b18287-s82的初始培养物的基因组或基因型中。在其它实施方式中,该edv可以包括直接或仅仅衍生自初始培养物的培养物,从而使得其基因组或基因型全部存在于系b18287-s82的初始培养物的基因组或基因型中。
7、将b18287-s82同核体与wbl-s290交配。应注意,该系是目前存在于株j15987中的亲代同核体之一(另一个是j11500-s80,其也用于本发明)。将所得的f1异核体株命名为j19109。
8、因此,本发明的另一个方面一般地涉及命名为wbl-s290的双孢蘑菇的新的独特的同核系,以及使用命名为wbl-s290的系的方法。在至少一个实施方式中,提供了包含双孢蘑菇系wbl-s290的至少一组染色体的培养物,其中所述至少一组染色体包含下表i中列出的序列表征的标志物基因座处的系wbl-s290的所有等位基因。如本文所公开的,双孢蘑菇系wbl-s290的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815northuniversity street,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68167。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。以适用的专利法要求的为准,在提交专利申请或颁发专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
9、在一个实施方式中,可以提供系wbl-s290的实质衍生品种(edv)。该edv可以包括衍生自初始培养物的培养物,其中所述初始培养物是系wbl-s290的培养物,从而使得其基因组或基因型的至少75%存在于系wbl-s290的初始培养物的基因组或基因型中。在其它实施方式中,其基因组或基因型的至少90%或95%或98%或99%存在于系wbl-s290的初始培养物的基因组或基因型中。在其它实施方式中,该edv可以包括直接或仅仅衍生自初始培养物的培养物,从而使得其基因组或基因型全部存在于系wbl-s290的初始培养物的基因组或基因型中。
10、本发明的另一个方面一般地涉及命名为j19109的新的独特的双孢蘑菇株培养物;它是经由两个同核培养物b18287-s82和wbl-s290的直接交配所获得的杂交株。株j19109的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815north university street,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68163。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。根据专利法,在提交优先权申请或颁发该株的专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
11、在一个实施方式中,可以提供株j19109的实质衍生品种(edv)。该edv可以包括衍生自初始培养物的双孢蘑菇的培养物,其中所述初始培养物是株j19109的培养物,从而使得其基因组或基因型的至少75%存在于株j19109的初始培养物的基因组或基因型中。作为另外一种选择,其它edv可以包括直接衍生自株j19109的初始培养物的培养物,从而使得其全部基因组或基因型存在于株j19109的初始培养物的基因组或基因型中。
12、重申一遍,将命名为b18287-s82的杂交蘑菇系的培养物(该系的代表性培养物已以nrrl保藏号68168保藏)与命名为wbl-s290的白色蘑菇系的培养物(该系的代表性培养物已以nrrl保藏号68167保藏)交配以获得命名为j19109的f1杂交蘑菇株(其代表性培养物已以nrrl保藏号68163保藏)。然后,将命名为j19109的f1株的培养物进行结实(收获,fruit)以从其获得同核孢子,由此获得同核系。然后,选择来自f1株j19109的同核系培养物。值得注意地,来自f1株j19109的同核系在染色体4、7和9上的系wbl-s290的着丝粒连锁基因座处缺少等位基因。
13、在从f1株j19109中选择同核系培养物后,将该培养物与命名为j11500-s80的蘑菇系的培养物交配,该蘑菇系被指出为目前存在于株j15987中的另一个亲代同核体。从该交配获得了所得的f2杂交蘑菇株,并对所得的f2杂交蘑菇株的培养物进行测试以确定ai性状的存在或不存在,其中在不存在ai性状的情况下,已从f2杂交蘑菇株中排除了ai性状。
14、因此,本发明的另一个方面一般地涉及命名为j11500-s80的双孢蘑菇的新的独特的同核系,以及使用命名为j11500-s80的系的方法。在至少一个实施方式中,提供了包含双孢蘑菇系j11500-s80的至少一组染色体的培养物,其中所述至少一组染色体包含下表i中列出的序列表征的标志物基因座处的系j11500-s80的所有等位基因。如本文所公开的,双孢蘑菇系j11500-s80的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815north university street,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68164。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。以适用的专利法要求的为准,在提交专利申请或颁发专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
15、在一个实施方式中,可以提供系j11500-s80的实质衍生品种(edv)。该edv可以包括衍生自初始培养物的培养物,其中所述初始培养物是系j11500-s80的培养物,从而使得其基因组或基因型的至少75%存在于系j11500-s80的初始培养物的基因组或基因型中。在其它实施方式中,其基因组或基因型的至少90%或95%或98%或99%存在于系j11500-s80的初始培养物的基因组或基因型中。在其它实施方式中,该edv可以包括直接或仅仅衍生自初始培养物的培养物,从而使得其基因组或基因型全部存在于系j11500-s80的初始培养物的基因组或基因型中。
16、将进一步理解,在一个或多个实施方式中,排除ai性状的f2杂交株将保留在株j15987中存在的至少两个有益性状。这些有益性状可以选自:与株j15987的菌盖形状一样圆的菌盖形状、与株j15987的菌柄一样粗的菌柄、与株j15987的菌肉一样厚的菌肉以及在l-a-b色彩测量标度上在株j15987的红色度(a)值的10%或5%或2.5%或1%或0.5%内的红色度(a)值。将理解术语“与……一样圆”和“与……一样粗”(“与……一样厚”)是指如所测量的,f2杂交株所结实蘑菇的菌盖、菌柄和/或菌肉在株j15987所结实蘑菇的菌盖、菌柄或菌肉的圆度或粗度(厚度)的统计学显著性范围内。在替代实施方式中,f2杂交株的蘑菇的菌盖形状的圆度可以在株j15987的蘑菇的菌盖形状的圆度的20%或15%或10%或5%内。在其它替代实施方式中,f2杂交株的蘑菇的菌柄的粗度可以在株j15987的蘑菇的菌柄的粗度的20%或15%或10%或5%内。在其它替代实施方式中,f2杂交株的蘑菇的菌肉的厚度可以在株j15987的蘑菇的菌肉的厚度的20%或15%或10%或5%内。
17、在一个实施方式中,选自f1株j19109的同核系可以是命名为j19109-s40的同核系。将理解将命名为j19109-s40的系培养物与命名为j11500-s80的蘑菇系交配,其中产生命名为j20176的f2杂交株。已发现杂交株j20176不具有ai性状。
18、因此,本发明的另一个方面一般地涉及命名为j19109-s40的双孢蘑菇的新的独特的同核系,以及使用命名为j19109-s40的系的方法。在至少一个实施方式中,提供了包含双孢蘑菇系j19109-s40的至少一组染色体的培养物,其中所述至少一组染色体包含下表i中列出的序列表征的标志物基因座处的系j19109-s40的所有等位基因。如本文所公开的,双孢蘑菇系j19109-s40的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815north university street,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68165。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。以适用的专利法要求的为准,在提交专利申请或颁发专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
19、在一个实施方式中,可以提供系j19109-s40的实质衍生品种(edv)。该edv可以包括衍生自初始培养物的培养物,其中所述初始培养物是系j19109-s40的培养物,从而使得其基因组或基因型的至少75%存在于系j19109-s40的初始培养物的基因组或基因型中。在其它实施方式中,其基因组或基因型的至少90%或95%或98%或99%存在于系j19109-s40的初始培养物的基因组或基因型中。在其它实施方式中,该edv可以包括直接或仅仅衍生自初始培养物的培养物,从而使得其基因组或基因型全部存在于系j19109-s40的初始培养物的基因组或基因型中。
20、本发明的另一个方面一般地涉及命名为j20176的新的独特的双孢蘑菇株培养物;它是经由两个同核培养物j11500-s80和j19109-s40的直接交配所获得的杂交株。株j20176的培养物的保藏已由农业研究机构培养物保藏中心(nrrl)(1815north universitystreet,peoria,illinois 61604usa)做出并接受。保藏日期为2022年6月10日。所保藏的培养物取自由sylvan,inc.,kittanning,pennsylvania,usa(登记受让人)在本发明申请提交日期之前保存的相同培养物。对保藏的所有限制都已取消,并且保藏旨在满足美国专利和商标局的所有保藏要求,包括37c.f.r.sec.1.801-1.809,以及布达佩斯条约规定的所有保藏要求。nrrl保藏号为68166。保藏将在保藏所保存30年,或在最后一次请求后保存5年,或在专利有效期内保存,以时间较长者为准,并将在此期间根据需要进行更换。根据专利法,在提交优先权申请或颁发该株的专利后,培养物将不可撤销地且无限制地或无条件地向公众发布。
21、在一个实施方式中,可以提供株j20176的实质衍生品种(edv)。该edv可以包括衍生自初始培养物的双孢蘑菇的培养物,其中所述初始培养物是株j20176的培养物,从而使得其基因组或基因型的至少75%存在于株j20176的初始培养物的基因组或基因型中。作为另外一种选择,其它edv可以包括直接衍生自株j20176的初始培养物的培养物,从而使得其全部基因组或基因型存在于株j20176的初始培养物的基因组或基因型中。
22、将进一步理解,在一个或多个实施方式中,命名为j20176的f2杂交株排除了ai性状,并且保留了在株j15987中存在的至少两个有益性状。这些有益性状可以选自:与株j15987的菌盖形状一样圆的菌盖形状、与株j15987的菌柄一样粗的菌柄、与株j15987的菌肉一样厚的菌肉以及在l-a-b色彩测量标度上在株j15987的红色度(a)值的10%或5%或2.5%或1%或0.5%内的红色度(a)值。将理解术语“与……一样圆”和“与……一样粗”(“与……一样厚”)是指f2杂交株j20176所结实蘑菇的菌盖、菌柄和/或菌肉在株j15987所结实蘑菇的菌盖、菌柄或菌肉的圆度或粗度(厚度)的统计学显著性范围内。在替代实施方式中,f2杂交株j20176的蘑菇的菌盖形状的圆度可以在株j15987的蘑菇的菌盖形状的圆度的20%或15%或10%或5%内。在其它替代实施方式中,f2杂交株j20176的蘑菇的菌柄的粗度(粗细度)可以在株j15987的蘑菇的菌柄的粗度的20%或15%或10%或5%内。在其它替代实施方式中,f2杂交株j20176的蘑菇的菌肉的厚度可以在株j15987的蘑菇的菌肉的厚度的20%或15%或10%或5%内。
23、可以看出本发明所述的方法有利地从双孢蘑菇株(至少包括株j20176)中排除了ai性状。更具体地,所述方法能够从源自(系出于)株j10165(也称为wbl)和株j11500的双孢蘑菇株排除攻击性不相容性(ai)性状(如美国专利no.9,622,428中所公开的),其中已知将命名为wbl-s290的白色蘑菇系的培养物与命名为j11500-s80的蘑菇系的培养物交配提供了具有ai性状的命名为j15987的杂交蘑菇株,该株的代表性培养物已以nrrl保藏号67646保藏。
24、相对于从来自f1株j19109的同核孢子获得同核系培养物,将理解当j19109异核体结实并收集f2孢子时,最初总计获得52个同核体,并且在下列表中的几个表中描述了这些中的16个。将全部52个同核体与j15987的第二亲代j11500-s80交配,并对所得菌株的商品化潜力进行广泛筛选。如下所示的所收集的数据表明j15987的有益性状得到了保持,并且更重要地,大多数新的j19109×j11500-s80杂交体不具有ai性状。选择一个杂交体(株j20176)进行受控测试。该菌株表现稳健,并因此是商业化的有力候选。
25、株j19109的同核系的整个池显示出与j19109-s40类似的潜力,并且全部被认为是有价值的育种系,其具有生产缺少ai性状的高质量蘑菇杂交体的潜力。
26、j19109衍生的杂交体的用途中的一个用途是生产用于销售的食用蘑菇作物。因此,从以上所指明的任何株系或菌株的培养物获得的蘑菇是本发明的一部分。另一个用途是通过菌株轮换(strain rotation)和“病毒破坏”效应来改善设施卫生。其它用途是将株j19109的基因材料掺入子代和衍生或后代培养物中,包括休眠和萌发的孢子和原生质体。如上所述,还存在其它用途。
27、真菌不寻常的生物学,并且具体地双孢蘑菇中的内源杂交,使得衍生具有基本相同表型的培养物的虚拟拷贝(虚拟复制物,virtual copy)变得容易和直接。在植物育种中,这些拷贝被称为实质衍生品种或edv。获得培养物(因此,根据定义,所述培养物是双孢蘑菇的单一初始培养物的edv)的方法包括(举例说明)体细胞选择、组织培养选择、单孢子萌发、多孢子萌发、自交、重复回交至初始培养物、诱变和转化。以下文献已报道了双孢蘑菇的dna-介导的转化:velcko,a.j.jr.,kerrigan,r.w.,macdonald,l.a.,wach,m.p.,schlagnhaufer,c.,and romaine,c.p.2004,expression of novel genes in agaricusbisporus using an agrobacterium-mediated transformationtechnique.mush.sci.16:591-597,和其中的参考文献,其作为参考并入本文。转化可以将单个新基因或等位基因引入初始培养物的基因组中。此外,最近关于其它密切相关的真菌的报道提出了通过crispr进行基因编辑的可能性。
28、edv是通过它们的基因型可明确辨别的,它们的基因型主要是或甚至完全是单一初始培养物的亚型(亚群,subset)。存在于双孢蘑菇edv中的初始基因型的百分比范围在100%或几乎100%(在单孢培养和体细胞选择的情况下),至99.x%(在通过dna介导的转化修饰的菌株的情况下),至90-99.x%(在一些单孢子或多孢子选择或一些诱变的情况下),至平均至少75-85%(在同胞子代交配(=自交)和回交到初始培养物的情况下)。多种基因型确定方法,包括以下所描述的方法和本领域熟知的其它方法,可以用于确定存在于另一培养物中的初始培养物的dna的百分比,并用于明确确定两种培养物之间的关系以及用于操纵或利用初始培养物的任何方法。
29、重复回交到初始培养物也产生了初始培养物的edv。在一个假设的实例中,在该过程的第一连续重复中,这一代的所得菌株将具有平均约75%的初始菌株的dna,而约25%的dna将由第二菌株或株系提供(贡献);当重复该过程时,初始菌株的dna表示(representation)将增加,在进一步3次连续重复后平均接近97%。通常,将理解与初始培养物具有75-100%基因型同一性的任何培养物都是初始培养物的edv的指示。还确立edv的edv也是初始株的edv。最后,由于双孢蘑菇在异核菌株和同核株系之间世代交替,因此实质衍生的标准同样适用于菌株和株系两者的培养物。
30、基因型指纹是对限定基因座处的基因型的描述,其中记录了所表征的等位基因的存在。这些指纹为识别克隆和初始菌株的edv的所有类型以及识别远交谱系中的祖先提供了强大且有效的技术。多种技术对于限定和表征(鉴定)基因型中的基因座和等位基因是可用的。全基因组测序(wgs)提供了最详细的方法,它允许对整个基因组的dna序列进行直接表征和比较。使用这种方法来产生引入了大量标志物基因座的稳健的基因型指纹,有可能在两个菌株之间建立(确立)关系性质,包括经过多个世代的系谱后代所关联的菌株。申请人已通过其菌株开发谱系的四至六代跟踪了遗传标志物。如果在初始菌株或株系中存在足够数目的独特标志物,则有可能在几个远交世代后鉴别来自初始菌株或株系的后代,而无需过度的实验。在一个假定实例中,在f4杂交体中,在4个远交世代后,初始单倍体株系的基因组表示的平均期望值为3.1%(50%/24),这对应于约1mb的双孢蘑菇的核基因组dna。基于申请人的分析,来自两个不相关的双孢蘑菇菌株中的每一个的dna的量通常可以含有约10000至约20000个单核苷酸多态性(snp),其中的任一个都可以提供将f4杂交体与初始株系相联系的区别标志物。通过使用多个独立的标志物,可以以非常高的成功概率和合理的置信度来鉴别菌株的祖先(原种,ancestor)。
31、具有生物学和商业价值(商业关注度)的一个性状是异核体不相容性(heterokaryon incompatibility)。这些自身/非自身识别系统的遗传学在担子菌类真菌,如伞菌属中尚未很好地阐明,但已知在其它真菌群中涉及多个独立基因座处的多个等位基因。注意,异核体不相容性发生在大多数真菌中,并且与本文所报道的攻击性不相容性状有很大不同。“正常的”不相容性通常是菌丝体相互作用的局部区域,其中两种基因型之间的生长减弱。据信该功能将防止病毒在同一物种的不同基因型之间转移。
32、应注意至少有两种影响蘑菇作物的双链rna病毒,即勒法郎士病(la francedisease)和蘑菇病毒x。异核体不相容性将阻止两个或更多个异核体的物理混合物中的成功接合和胞质连续性,借此产生“病毒破坏(virus-break)”。在蘑菇农场,通过用掺入与初始菌株不相容的另一不同菌株的接种物和作物材料替换掺入初始菌株的作物材料(堆肥、菌种体、覆盖接种物)来进行病毒破坏。在病毒破坏方法的最有效的实施中,蘑菇农场的初始株的所有生物材料被第二不相容菌株的生物材料替换。菌株不相容性为病毒从设施内的生物库向新作物的移动产生了有效(如果不是绝对)的屏障。具有不同基因型的蘑菇株之间的轮转栽培使用也可以中断外源害虫和病原体的感染和侵染循环。
33、如以上所提及的,杂交蘑菇株生产者一直在寻找能让种植者成功且能获利地生产作物的杂交株。在衍生自j19109的新杂交体及其衍生株或后代株的情况下,迄今为止所记录的积极属性包括有吸引力的外观(与市场领先者a-15相比,圆形菌盖、粗菌柄、良好的抗压伤性(bruise resistance),所有这些都对消费者有吸引力),以及可以匹配或超过诸如a-15的菌株的总产量。在我们的实例中,蘑菇种植者可以通过使用本文所描述的新杂交体获得高质量蘑菇的高产作物。另外,这些菌株与j15987的高质量相匹配,j15987是本发明申请人所开发的早期菌株,具有缺少攻击性不相容性表型的显著优势。
34、目前可用于蘑菇产业的菌株使种植者能够成功生产蘑菇作物,并且通常是有利可图的。有几个因素会影响所实现的成功和盈利程度。例如,相对于目前上市和商业种植的菌株,菌株必须能够在两到三批暴发或两到三次出菇潮中产生至少相当的作物产量。此外,所生产的蘑菇的一些物理性质,例如,菌盖颜色类别,以及常规大小和尺寸,如菌盖直径,允许蘑菇以熟悉的产品类别上市。
35、j19109 ssi×j11500-s80杂交体能满足那些市场需求,并解决了目前市场上无法获得这种菌株的问题。另外,当与其它现有的白色栽培种相比时,这组新的菌株在基因型上不同(参见下表i和ii),这解决了与对单一栽培作物的依赖性有关的潜在问题。
36、本发明的一个或多个方面可以通过由本发明所开发的双孢蘑菇的杂交蘑菇培养物来实现。因此,本发明涵盖了掺入上述株系或菌株的任何培养物的产品。这些产品可以包括菌丝体(mycelium)、菌种体(spawn)、新鲜或加工的蘑菇、蘑菇孢子、蘑菇菌种体、蘑菇制备物(mushroom preparations)和提取物和级分(fractions)、蘑菇块(mushroom pieces)、蘑菇接种物、覆盖接种物(casing inoculum)、覆盖菌种体(casing spawn)、覆盖土壤、接种的堆肥、定殖的堆肥、后耕作的堆肥(收割后的堆肥,post-cropped compost)和松散颗粒物(friable particulate matter)。
37、此外,上述任何株系或菌株的培养物的多个部分可能具有价值和商业应用。因此,设想了选自菌丝、蘑菇、孢子、细胞、核和原生质体的部分。更进一步,在一个或多个实施方式中,设想了单细胞微生物。因此,提供了上述株系或菌株的任何培养物的细胞。
38、应注意已保藏了株j20176(已以nrrl保藏号68166保藏了该菌株的代表性培养物)以提供满足本发明要求的可能株系的适合实例。因此,株j20176可以包括培养物的多个部分,包括菌丝、孢子以及细胞和细胞部分,包括核、线粒体、细胞质、原生质体、dna、rna、蛋白质、细胞膜和细胞壁,所述部分存在于培养物的营养菌丝体中和通过所述培养物所产生的蘑菇中。所述孢子可以是休眠的或萌发的孢子,并且可以包括掺入其中的异核体和同核体。
39、可以产生一种或多种掺入了指明为j19109的后代菌株的双孢蘑菇的杂交蘑菇培养物的产品。这些产品包括菌丝体、菌种体、接种物、覆盖接种物、新鲜蘑菇、加工蘑菇、蘑菇提取物和级分、蘑菇块和选自谷物、堆肥和松散颗粒物的定殖基质。将理解蘑菇块是指蘑菇本身的菌柄、菌盖和其它较大的部分。
40、本发明的一个或多个其它方面可以通过菌株,如j19109的杂交蘑菇培养物的实质衍生品种(edv)来实现。在一个或多个实施方式中,通过实质衍生所产生的双孢蘑菇培养物具有j19109衍生交配体的基本特征中的至少一种。对于本发明,作为本文所描述的本发明的直接结果的优良菌株的实例,我们保藏了一个交配体,j19109-s40×j11500-s80,其被称为j20176。j20176是缺少ai性状的菌株,并且相对于u1 edv,如a-15,具有j15987的菌盖圆度、菌肉厚度、产量表现和产量时间等其它特征。
41、本发明的其它方面可以通过用于产生双孢蘑菇杂交培养物的方法来实现,该方法包括将同核体,如j19109-s40(以nrrl保藏号68165保藏)与第二同核体交配的步骤。在一个实施方式中,第二同核系j11500-s80,其培养物以nrrl保藏号68164保藏。这种交配提供了蘑菇培养物j20176,其证实对一组基于u1 edv的商品化栽培种,例如a-15的正常拮抗模式。这种拮抗作用证实了株j20176的遗传独特性。在一个或多个实施方式中,所述方法进一步包括提供本发明的蘑菇培养物,所述培养物由菌丝体、菌种体、接种物、覆盖接种物、新鲜蘑菇、加工蘑菇、蘑菇部分、蘑菇提取物和级分、蘑菇块和选自谷物、堆肥和松散颗粒物的定殖基质组成。在其它实施方式中,所述方法可以包括在衍生培养物中提供蘑菇培养物,所述衍生培养物选自同核体、异核体、非整倍体、体细胞继代培养物、组织外植体培养物、原生质体、休眠孢子、萌发孢子、近交后代和远交后代、转基因培养物、基因编辑培养物和具有单基因座转换的基因组的培养物。
42、本发明的其它方面可以通过具有基因型指纹的双孢蘑菇的杂交蘑菇培养物来实现,所述基因型指纹在标志物基因座its、p1n150-g3-2、mfpc-1-elf、an、af和ff处具有特征。在一个或多个实施方式中,所述培养物具有在标志物基因座处具有表ii中所描述的特征的基因型指纹,其中所述指纹的所有特征都存在于基因型指纹中。需要明确的是,减数分裂和随机分配(random assortment)将确保j19109后代的每个同核孢子将具有独特的标志物模式。基于所遗传的dna标志物,本领域技术人员将直接推断出j19109同核体育种原种(育种储库,breeding stocks)是后代。
43、本发明的一个或多个其它方面可以通过由以上方法所产生的培养物、细胞或包括所述细胞的培养物来实现。因此,一个或多个实施方式可以包括进一步包括生长杂交蘑菇培养物以产生杂交蘑菇和蘑菇部分的步骤的方法。其它实施方式可以提供这类方法,其中所产生的杂交蘑菇培养物或细胞包括在标志物基因座its、p1n150-g3-2、mfpc-1-elf、an、af和ff处具有特征的标志物谱,其中所述标志物谱的所有特征也存在于株j20176和j19109的标志物谱中。仍有其它实施方式可以提供这类方法,其中所产生的杂交蘑菇培养物或细胞包括在表i中所描述的标志物基因座处具有特征的标志物谱,其中所述标志物谱的所有特征也存在于株j20176的标志物谱中。
44、本发明相对于已有的涉及双孢蘑菇和培养物的现有技术的这些和其它优势将根据以下描述而变得显而易见,并通过下文描述和主张的本发明来实现。
技术研发人员:迈克尔·凯斯勒,米歇尔·舒尔茨,马克·洛夫特斯,阿尼萨·阿米尼,西尔维·德尔贝克,韦斯·舒尔茨,理查德·克里根,马克·瓦赫
技术所有人:施尔丰公司
备 注:该技术已申请专利,仅供学习研究,如用于商业用途,请联系技术所有人。
声 明 :此信息收集于网络,如果你是此专利的发明人不想本网站收录此信息请联系我们,我们会在第一时间删除